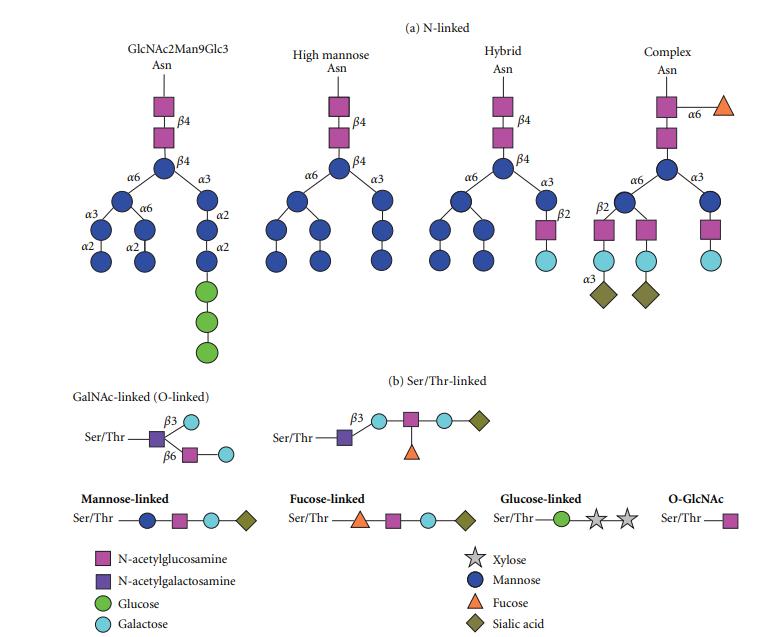
Whole cell lysate (20 μl) was combined with 80 μl of 0.05 N H2SO4 (hydrolysis reagent) and incubated at 80°C for 60 min. When you loved this informative article and you would like to receive more info regarding china n-acetylneuraminic acid powder generously visit our own website. Samples were briefly centrifuged at 14,000 revolution/min (16,000 g), after which 20 μl of 1 M NaOH (neutralization reagent) was added and the mixture centrifuged again at 14,000 revolution/min. Afterwards, wells were washed 3 times, 100 µl of substrate solution was added for 30 min at RT and the optical density (OD) was measured at 405 nm by an ELISA reader (TECAN Infinite 200 PRO). The wells were washed 3 times and then 100 µl of a specific alkaline phosphatase labelled antibody for terminal complement complex (TCC, C5b-9; provided by the manufacturer) was added and incubated for 30 min at RT. Finally, we demonstrate the contribution of terminal sialic acids to endothelial barrier integrity. Additionally, it will be important to determine whether acetylated sialic acids or (2,8) dimeric-linked sialic acids play a key role in determining barrier integrity. Two components of glycocalyx structure that may play a role in determining function in glycoproteins are 1) the terminal sugar residue(s) on the oligosaccharide chain(s) and 2) the amino acid residue attachment site (N- vs. Neuraminidases are enzymes that cleave via hydrolysis α(2-3)-, α(2-6)-, and α(2-8)-linked terminal sialic acid residues bound to Gal, GlcNac, GalNAc, AcNeu, or GlyNeu residues of oligosaccharides, glycolipids, and glycoproteins (17). Neuraminidases from different sources exhibit different specificities for sialic acid linkages hydrolyzed (4, 24). The lectin from Arachis hypogaea binds to the sequence Gal(β1,3)GalNAc, also known as T-antigen (19, 24). When the T-antigen sequence is sialylated, lectin from Arachis hypogaea does not bind to the disaccharide (10). However, as in the case of red blood cells, following treatment with neuraminidase, the T-antigen is exposed on the cell surface allowing the lectin to bind (19). Indeed, this approach has already been used to demonstrate loss of sialic acids from pulmonary endothelial cell surfaces (26). For these experiments, PAECs and PMVECs were treated with neuraminidase from Clostridium perfringens, which cleaves α(2-3)-, α(2-6)-, and α(2-8)-terminal sialic acid residues (3, 4, 17). The Arachis hypogaea lectin did not bind to control cells but exhibited strong binding to neuraminidase-treated cells as evidenced by positive fluorescence in treated cells (Fig. 2B), revealing the underlying Gal(β1,3)GalNAc epitope.
 "conservative amino acid substitutions," in one or a few amino acids in an amino acid sequence are substituted with different amino acids with highly similar properties are also readily identified as being highly similar to a particular amino acid sequence, or to a particular nucleic acid sequence which encodes an amino acid. In the present study, we quantitate sialic acids present in PAECs and PMVECs and utilize exoglycosidase enzymes and stereospecific fluorescent lectin binding to identify specific sialic acid configurations on the two cell types. Cultured monolayers of PAECs or PMVECs were washed twice with HBSS, following which neuraminidase solutions were added. Lectin solutions were prepared at the following concentrations in HBSS: TRITC-tagged Arachis hypogaea agglutinin (10 μg/ml), FITC-tagged Maackia amurensis agglutinin (MAA, 20 μg/ml), Texas Red-tagged Sambucus nigra agglutinin (SNA, 20 μg/ml), or FITC-tagged SNA (10 μg/ml). First, within 2 h of either neuraminidase treatment, the α(2,6)-linked sialic acids in PAECs were hydrolyzed as evidenced by loss of FITC-tagged SNA fluorescence (Fig. 5A). Second, we observed overall disruption of the PAEC monolayer following treatment with either neuraminidase although there were characteristic differences in what the resultant disrupted monolayer looked like.
"conservative amino acid substitutions," in one or a few amino acids in an amino acid sequence are substituted with different amino acids with highly similar properties are also readily identified as being highly similar to a particular amino acid sequence, or to a particular nucleic acid sequence which encodes an amino acid. In the present study, we quantitate sialic acids present in PAECs and PMVECs and utilize exoglycosidase enzymes and stereospecific fluorescent lectin binding to identify specific sialic acid configurations on the two cell types. Cultured monolayers of PAECs or PMVECs were washed twice with HBSS, following which neuraminidase solutions were added. Lectin solutions were prepared at the following concentrations in HBSS: TRITC-tagged Arachis hypogaea agglutinin (10 μg/ml), FITC-tagged Maackia amurensis agglutinin (MAA, 20 μg/ml), Texas Red-tagged Sambucus nigra agglutinin (SNA, 20 μg/ml), or FITC-tagged SNA (10 μg/ml). First, within 2 h of either neuraminidase treatment, the α(2,6)-linked sialic acids in PAECs were hydrolyzed as evidenced by loss of FITC-tagged SNA fluorescence (Fig. 5A). Second, we observed overall disruption of the PAEC monolayer following treatment with either neuraminidase although there were characteristic differences in what the resultant disrupted monolayer looked like.
Consistent with this published study, our results demonstrated that transduction with AAV1 and AAV6 was not inhibited by mucin (Fig. (Fig.9).9). The relative transduction efficiency was determined by comparison with control untreated cells and is presented in log scale. These data suggested that AAV1 and AAV6 use N-linked sialic acid for efficient transduction. In addition, many bacteria including E. coli K12 are able to catabolise Neu5Ac and use it as a carbon energy source. We initially wished to determine whether sialic acids are expressed in the glycocalyx of PAECs and PMVECs. Neuraminidase treatment of PAECs and PMVECs. Pulmonary artery endothelial cells (PAECs) and pulmonary microvascular endothelial cells (PMVECs) express sialic acids. Cells were grown to confluence in eight-well ECIS arrays. Electric cell-substrate impedance sensing (ECIS) experiments were conducted using an Applied Biophysics Model 1600R instrument (Applied Biophysics, Troy, NY). The standard curve was prepared using trypsin with the lowest concentration at 1 ng/ml.
 An alternative is the enzymatic synthesis of Neu5Ac from N-acetylmannosamine (ManNAc) and pyruvate using the N-acetylneuraminic acid aldolase. Dotted lines represent the enzymatic reactions that have been abolished to make possible the production of Neu5Ac by bacterial fermentation. We control the quality very well all the time, every batch of the powder must have the test data before finished in package. The cut-off point for positive staining was above the level of the control isotype mAbs. This was followed by the addition of neuraminidase solution or buffer control to each well and continued resistance measurement 4 (or more) h. Hank's Balanced Salt Solution (HBSS) was purchased from Invitrogen (Carlsbad, CA). Perfusion was maintained at a constant flow (0.045 ml/g body wt) with Earle's buffered solution containing 4% purified bovine serum albumin and calcium chloride adjusted at 5.5 mM. After completion of the second Kf, venous pressure was returned to baseline for 5 min, and the lower right lobe was immersion fixed in buffered formalin.
An alternative is the enzymatic synthesis of Neu5Ac from N-acetylmannosamine (ManNAc) and pyruvate using the N-acetylneuraminic acid aldolase. Dotted lines represent the enzymatic reactions that have been abolished to make possible the production of Neu5Ac by bacterial fermentation. We control the quality very well all the time, every batch of the powder must have the test data before finished in package. The cut-off point for positive staining was above the level of the control isotype mAbs. This was followed by the addition of neuraminidase solution or buffer control to each well and continued resistance measurement 4 (or more) h. Hank's Balanced Salt Solution (HBSS) was purchased from Invitrogen (Carlsbad, CA). Perfusion was maintained at a constant flow (0.045 ml/g body wt) with Earle's buffered solution containing 4% purified bovine serum albumin and calcium chloride adjusted at 5.5 mM. After completion of the second Kf, venous pressure was returned to baseline for 5 min, and the lower right lobe was immersion fixed in buffered formalin.